
| Website | Home URL |
| Division | Biochemistry |
| Title |
|
| Specialties |
|
| cfclarke@g.ucla.edu | |
| Office | Young Hall 5072B |
| Office Phone | (310) 825-0771 |
| Lab | Young Hall 2107, 5073 |
| Office Lab | (310) 825-0668 |
Short Biography
Clarke joined the chemistry and biochemistry department at University of California, Los Angeles in 1993. Her research considers how cells synthesize coenzyme Q, a non-protein chemical compound that is also known as ubiquinone. Further, she is also interested in studying the aging process and the impact of genetic mutations, utilizing our understandings of diet, environment and genotype to determine longevity.
In June 2016, Clarke became the first woman to lead the UCLA Department of Chemistry and Biochemistry. Clarke was appointed Dean of Special Projects in the UCLA Division of Physical Sciences in 2019.
Research Interests
Introduction – Ubiquinone (coenzyme Q or Q) functions in cells as a redox-active coenzyme of mitochondrial and plasma membrane electron transport, as well as an essential lipid soluble antioxidant. Human dietary supplementation with Q appears to have beneficial effects in slowing the progression of neuro- and muscle-degenerative diseases. However, Q is also involved in generating reactive oxygen species, through the adventitious reduction of dioxygen to superoxide by the ubisemiquinone radical, normally generated during mitochondrial electron transport. Thus, it is not clear how dietary supplementation with Q impacts the Dr. Jekyll/Mr. Hyde aspects of Q function. Cells are capable of synthesizing Q, but much remains to be learned about the sites of its synthesis, mechanisms of inter- and intra-cellular transport, and the regulation and enzymology of its biosynthesis. Research in my laboratory identified eight of the eleven polypeptides required for Q biosynthesis. The goals of my research are to characterize the Coq polypeptides responsible for production of Q and to determine how their activity can be modulated for optimal health.
Research Overview –
(1) We discovered a new Q biosynthetic pathway and showed that yeast are able to synthesize Q from a new aromatic ring precursor, para-aminobenzoic acid (pABA) [77]. This pathway operates in addition to the classic Q biosynthetic pathway that emanates from 4-hydroxybenzoic acid (4HB). We suggest a mechanism where Schiff base mediated deimination forms DMQ6 quinone, thereby eliminating the nitrogen contributed by pABA. This scheme results in the convergence of the 4HB and pABA pathways in eukaryotic Q biosynthesis and has implications regarding the action of pABA-based antifolates. This work is funded by a grant from NSF.
(2) The findings we made in the yeast model have shed much light on diseases resulting from Q deficiency [82, 85]. We showed that human Coq8 and Coq6 polypeptide homologues function to rescue the respective yeast coq mutants [80, 81]. We discovered that yeast Coq8p, and its human homologue ADCK3 function in the phosphorylation of Coq3p, Coq5p, and Coq7p [80]. The kinase function of Coq8p is proposed to be important in the maintenance or formation of a high molecular mass Coq polypeptide complex, which is required for Q biosynthesis.
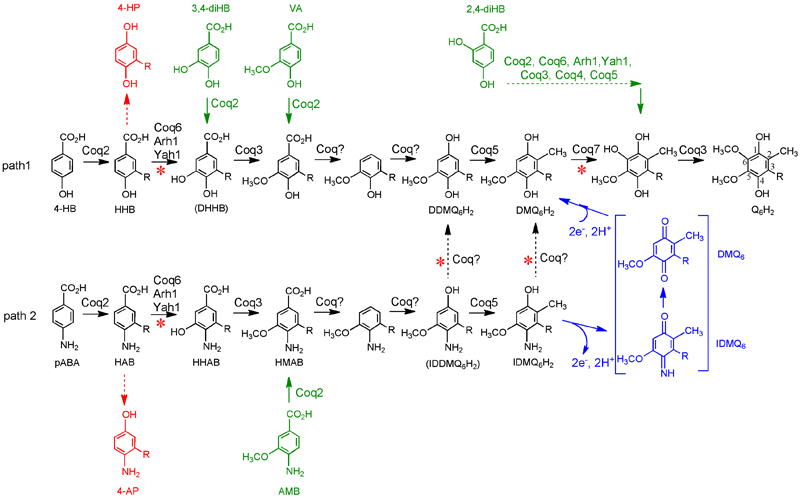
(3) Over-expression of Coq8p in many of the yeast coq null mutants restores steady state levels of Coq polypeptides. This has been a particularly important discovery, because the yeast coq null mutants (coq3-coq9) previously accumulated the same early Q-intermediate. Coq8 over-expression has enabled us to detect late-stage Q-intermediates that were previously elusive [86]. Finally, we used synthetic analogues of 4-hydroxybenzoic acid to bypass deficient biosynthetic steps and showed that 2,4-dihydroxybenzoic acid is able to restore Q6 biosynthesis and respiratory growth in a coq7 null mutant over-expressing Coq8. The over-expression of Coq8 and the use of analogues of 4-HB represent new tools to elucidate the Q biosynthetic pathway. These recent discoveries are depicted in Figure 1.
 |
|
Figure 1. S. cerevisiae Q6 biosynthetic pathway: accumulation of Q6 biosynthetic intermediates caused by the over-expression of Coq8 in Dcoq strains. The classic Q biosynthetic pathway is shown in path 1 emanating from 4-hydroxybenzoic acid (4-HB). Coq1 (not shown) synthesizes the hexaprenyl-diphosphate tail which is transferred by Coq2 to 4-HB to form 3-hexaprenyl-4-hydroxybenzoic acid (HHB). R represents the hexaprenyl tail present in all intermediates from HHB to Q6. Alternatively, path 2 shows para-aminobenzoic acid (pABA) is prenylated by Coq2 to form 3-hexaprenyl-4-aminobenzoic acid (HAB). Both HHB and HAB are early Q-intermediates, readily detected in each of the coq null strains (Dcoq3-Dcoq9). The numbering of the aromatic carbon atoms used throughout this study is shown on the reduced form of Q6, Q6H2. Coq8 over-expression in certain Dcoq strains leads to the accumulation of the following compounds: 4-AP, 3-hexaprenyl-4-aminophenol; 4-HP, 3-hexaprenyl-4-hydroxyphenol; HHAB, 3-hexaprenyl-4-amino-5-hydroxybenzoic acid; HMAB, 3-hexaprenyl-4-amino-5-methoxybenzoic acid; DDMQ6, the oxidized form of demethyl-demethoxy-Q6H2; IDMQ6, 4-imino-demethoxy-Q6; DMQ6H2, demethoxy-Q6H2. IDDMQ6H2, 2-demethyl-4-amino-demethoxy-Q6H2 and DHHB, 3-hexaprenyl-4,5-dihydroxybenzoic acid are shown but have not been detected in this study. Black dotted arrows (from path 2 to path 1) designate the replacement of the C4-amine with a C4-hydroxyl and correspond to the C4-deamination reaction. A putative mechanism to replace the C4-imino group with the C4-hydroxy group is shown in bracket in blue on IDMQ6 but could also occur on IDDMQ6 (not shown). 4-AP and 4-HP which are formed upon inhibition of the C5-hydroxylation catalyzed by Coq6 are shown in red. Analogues of 4-HB and pABA allowing the bypass of certain steps in Q biosynthesis are indicated in green. Steps defective in the Dcoq9 strain are designated with a red asterisk. |
4) In collaboration with Retrotope, we have employed novel isotope-reinforced polyunsaturated fatty acids (PUFAs) that reveal the importance of Q as an essential antioxidant [79, 87]. Polyunsaturated fatty acids (PUFA) undergo autoxidation and generate reactive carbonyl compounds that are toxic to cells and associated with apoptotic cell death, age-related neurodegenerative diseases, and atherosclerosis. PUFA autoxidation is initiated by the abstraction of bis-allylic hydrogen atoms. Replacement of the bis-allylic hydrogen atoms with deuterium atoms (termed site-specific isotope-reinforcement) arrests PUFA autoxidation due to the isotope effect. We investigate the effects of different isotope-reinforced PUFAs and natural PUFAs on autoxidation kinetics, and on the viability of coenzyme Q-deficient Saccharomyces cerevisiae coq mutants and wild-type yeast subjected to copper stress. Cells treated with a C11-BODIPY fluorescent probe to monitor lipid oxidation products show that lipid peroxidation precedes the loss of viability due to H-PUFA toxicity. We show that replacement of just one bis-allylic hydrogen atom with deuterium is sufficient to arrest lipid autoxidation. In contrast, PUFA reinforced with two deuterium atoms at mono-allylic sites remain susceptible to autoxidation. Surprisingly, yeast treated with a mixture of approximately 20%:80% isotope-reinforced D-PUFA: natural H-PUFA are profoundly protected from lipid autoxidation-mediated cell killing. Our findings show that inclusion of only a small fraction of PUFA deuterated at the bis-allylic sites is sufficient to profoundly inhibit the chain reaction of non-deuterated PUFA.
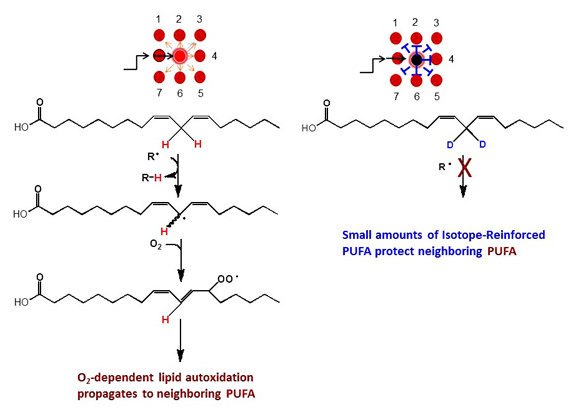
Fig. 2. Replacement of bis-allylic H atoms with deuterium (D) arrests autoxidation of PUFA. A theoretical chain reaction is depicted where a single initiation event producing a lipid peroxyl radical (denoted by –OO•) starts a chain reaction of lipid autoxidation that in the presence of O2, may continue indefinitely and produce many molecules of lipid peroxides; susceptible phospholipid molecules containing a PUFA acyl chain are designated by a red dot. The presence of 20% isotope-reinforced PUFA (denoted by a black dot) inhibit (or slow) chain propagation. Propagation is inhibited for PUFA neighboring the D-PUFA.
Honors & Awards
2001 Ellison Medical Foundation Senior Scholar Award, Ellison Medical Foundation
2009 Hanson-Dow Award for Excellence in Teaching, UCLA
2018 WHS Hall of Fame, Whittier High School
2019 BSF Research Grant Award, BSF
2021 Career Development Award
Representative Publications
Below is the list of the most recent publications. For the full list, please visit here.
127. Feustel, K. J.; Clarke,* C. F. | Resurrecting an ancient coenzyme Q metabolon – Nat. Catal. 2024
126. Alessandra Feoli, Giulia Iannelli, Alessandra Cipriano, Ciro Milite, Lei Shen, Zhihao Wang, Andrea Hadjikyriacou, Troy L. Lowe, Cyrus Safaeipour, Monica Viviano, Giuliana Sarno, Elva Morretta, Maria Chiara Monti, Yanzhong Yang, Steven G. Clarke*, Sandro Cosconati, Sabrina Castellano, and Gianluca Sbardella. Identification of a Protein Arginine Methyltransferase 7 (PRMT7)/Protein Arginine Methyltransferase 9 (PRMT9) Inhibitor – J. Med. Chem. 2023
125. Guile, M. D., Jain, A., Anderson, K. A., Clarke, C. F. (2023) “New Insights on the Uptake and Trafficking of Coenzyme Q.” Antioxidants 12, 1391.
124. Wang, S., Jain, A., Novales, N. A., Nashner, A. N., Tran, F., Clarke, C. F. (2022) “Predicting and Understanding the Pathology of Single Nucleotide Variants in Human COQ Genes.” Antioxidants 11, 2308.
123. Yang, M. L., Connolly, S. E., Gee, R. J., Lam, T. T., Kanyo, J., Peng, J., Guyer, P., Syed, F., Tse, J. M., CLarke, S. G., Clarke, C. F., James, E. A., Speake, C., Evans-Molina, C., Arvan, P., Herold, K. C., Wen, L., Mamula, M. J. (2022) “Carbonyl Posttranslational Modification Associated with Early-Onset Type 1 Diabetes Autoimmunity” Diabetes 71, 1979-1993.
122. Latimer, S., Keene, S. A>, Stutts, L. R., Berger, A., Bernert, A. C., Soubeyrand, E., Wright, J., Clarke, C. F., Block, A. K., Colquhoun, T. A., Elowsky, C., Christensen, A., Wilson, M. A., Basset, G. J. (2021) “A dedicated flavin-dependent monooxygenase catalyzes the hydroxylation of demethoxyubiquinone into ubiquinone (coenzyme Q) in Arabidopsis” J Biol Chem 297, 101283.
121. Ayer, A., Fazakerley, D. J., Suarna, C., Maghzal, G. J., Sheipouri, D., Lee, K. J., Bradley, M. C., Fernández-del-Río, L., Tumanov, S., Kong, S. M., van der Veen, J. N., Yang, A., Ho, J. W. K., Clarke, S. G., James, D. E., Dawes, I. W., Vance, D. E., Clarke, C. F., Jacobs, R. L., Stocker, R. (2021) “Genetic screening reveals phospholipid metabolism as a key regulator of the biosynthesis of the redox-active lipid coenzyme Q.” Redox Biol 46, 102127.
120. Fernández-del-Río, L., Rodriguez-Lopez, S., Gutierrez-Casado, E., Gonzalez-Reyes, J. A., Clarke, C. F., Buron, M. I., and Villalba, J. M. (2021) “Regulation of hepatic coenzyme Q biosynthesis by dietary omega-3 polyunsaturated fatty acids.” Redox Biol 46, 102061.
119. Fernández-del-Río, L., and Clarke, C. F. (2021) “Coenzyme Q Biosynthesis: An Update on the Origins of the Benzenoid Ring and Discovery of New Ring Precursors.” Metabolites 11, 385-401.
118. Acoba, M. G., Alpergin, E. S. S., Renuse, S., Fernández-del-Río, L., Lu, Y. W., Khalimonchuk, O., Clarke, C. F., Pandey, A., Wolfgang, M. J., and Claypool, S. M. (2021) “The mitochondrial carrier SFXN1 is critical for complex III integrity and cellular metabolism.” Cell Rep 34, 108869.
117. Fernández-del-Río, L., Soubeyrand, E., Basset, G. J., and Clarke, C. F. (2020) “Metabolism of the Flavonol Kaempferol in Kidney Cells Liberates the B-ring to Enter Coenzyme Q Biosynthesis.” Molecules 25, 2955-2962.
116. Awad, A. M., Nag, A., Pham, N. V. B., Bradley, M. C., Jabassini, N., Nathaniel, J., and Clarke, C. F. (2020) “Intragenic suppressor mutations of the COQ8 protein kinase homolog restore coenzyme Q biosynthesis and function in Saccharomyces cerevisiae.” PLoS One 15, e0234192.
115. Fernández-del-Río, L., Kelly, M. E., Contreras, J., Bradley, M. C., James, A. M., Murphy, M. P., Payne, G. S., and Clarke, C. F. (2020) “Genes and lipids that impact uptake and assimilation of exogenous coenzyme Q in Saccharomyces cerevisiae.” Free Radic Biol Med 154, 105-118.
114. Widmeier, E., Yu, S., Nag, A., Chung, Y. W., Nakayama, M., Fernández-Del-Río, L., Hugo, H., Schapiro, D., Buerger, F., Choi, W. I., Helmstadter, M., Kim, J. W., Ryu, J. H., Lee, M. G., Clarke, C. F., Hildebrandt, F., and Gee, H. Y. (2020) “ADCK4 Deficiency Destabilizes the Coenzyme Q Complex, Which Is Rescued by 2,4-Dihydroxybenzoic Acid Treatment.” J Am Soc Nephrol 31, 1191-1211.
113. Bradley, M. C., Yang, K., Fernández-Del-Río, L., Ngo, J., Ayer, A., Tsui, H. S., Novales, N. A., Stocker, R., Shirihai, O. S., Barros, M. H., and Clarke, C. F. (2020) “COQ11 deletion mitigates respiratory deficiency caused by mutations in the gene encoding the coenzyme Q chaperone protein Coq10.” J Biol Chem 295, 6023-6042.
112. Bernert, A. C., Jacobs, E. J., Reinl, S. R., Choi, C. C. Y., Roberts Buceta, P. M., Culver, J. C., Goodspeed, C. R., Bradley, M. C., Clarke, C. F., Basset, G. J., and Shepherd, J. N. (2019) “Recombinant RquA catalyzes the in vivo conversion of ubiquinone to rhodoquinone in Escherichia coli and Saccharomyces cerevisiae.” Biochim Biophys Acta Mol Cell Biol Lipids 1864, 1226-1234.
111. Tsui, H. S., Pham, N. V. B., Amer, B. R., Bradley, M. C., Gosschalk, J. E., Gallagher-Jones, M., Ibarra, H., Clubb, R. T., Blaby-Haas, C. E., and Clarke, C. F. (2019) “Human COQ10A and COQ10B are distinct lipid-binding START domain proteins required for coenzyme Q function.” Journal of lipid research 60, 1293-1310.
110. Eisenberg-Bord, M., Tsui, H. S., Antunes, D., Fernández-Del-Río, L., Bradley, M. C., Dunn, C. D., Nguyen, T. P. T., Rapaport, D., Clarke, C. F., and Schuldiner, M. (2019) “The Endoplasmic Reticulum-Mitochondria Encounter Structure Complex Coordinates Coenzyme Q Biosynthesis.” Contact (Thousand Oaks) 2, 1-14.
109. Widmeier, E., Airik, M., Hugo, H., Schapiro, D., Wedel, J., Ghosh, C. C., Nakayama, M., Schneider, R., Awad, A. M., Nag, A., Cho, J., Schueler, M., Clarke, C. F., Airik, R., and Hildebrandt, F. (2019) “Treatment with 2,4-Dihydroxybenzoic Acid Prevents FSGS Progression and Renal Fibrosis in Podocyte-Specific Coq6 Knockout Mice.” J Am Soc Nephrol 30, 393-405.
108. Awad, A. M., Bradley, M. C., Fernández-Del-Río, L., Nag, A., Tsui, H. S., and Clarke, C. F. (2018) “Coenzyme Q10 deficiencies: pathways in yeast and humans.“ Essays Biochem 62, 361-376.
107. Awad, A. M., Venkataramanan, S., Nag, A., Galivanche, A. R., Bradley, M. C., Neves, L., Douglass, S., Clarke, C. F., Johnson T. L. (2017) “Chromatin-remodeling SWI/SNF complex regulates coenzyme Q6 synthesis and a metabolic shift to respiration in yeast.” J Biol Chem 292, 14851-14866.
106. He, C. H., Black, D. S., Allan, C. M., Meunier, B., Rahman, S., Clarke, C. F. (2017) Human COQ9 rescues a coq9 yeast mutant by enhancing coenzyme Q biosynthesis from 4-hydroxybenzoic acid and stabilizing the CoQ synthome. Frontiers in Physiology 8, 463.
105. Fern�ndez-del-Rio, L., Nag, A., Guti�rrez Casado, E., Ariza, J., Awad, A. M., Joseph, A. I., Kwon, O., Verdin, E., de Cabo, R., Schneider, C., Torres, J. Z., Bur�n, M. I., Clarke, C. F., Villalba, J. M. (2017) “Kaempferol increases levels of coenzyme Q in kidney cells and serves as a biosynthetic ring precursor.” Free Rad Biol Med 110, 176-187.
104. Clarke CF, Allan CM. (2015) “Biochemistry: Unexpected role for vitamin B2.” Nature. 2015 Jun 25;522(7557):427-8. doi: 10.1038/nature14536. Epub 2015 Jun 17. No abstract available. PMID: 26083748
103. He CH, Black DS, Nguyen TP, Wang C, Srinivasan C, Clarke CF. (2015) “Yeast Coq9 controls deamination of coenzyme Q intermediates that derive from para-aminobenzoic acid.” Biochim Biophys Acta. 2015 Sep;1851(9):1227-39. doi: 10.1016/j.bbalip.2015.05.003. Epub 2015 May 23. PMID: 26008578
102. Xie LX, Williams KJ, He CH, Weng E, Khong S, Rose TE, Kwon O, Bensinger SJ, Marbois BN, Clarke CF. (2015) “Resveratrol and para-coumarate serve as ring precursors for coenzyme Q biosynthesis.” J Lipid Res. 2015 Apr;56(4):909-19. doi: 10.1194/jlr.M057919. Epub 2015 Feb 14. PMID: 25681964
101. Allan CM, Awad AM, Johnson JS, Shirasaki DI, Wang C, Blaby-Haas CE, Merchant SS, Loo JA, Clarke CF. (2015) “Identification of Coq11, a new coenzyme Q biosynthetic protein in the CoQ-synthome in Saccharomyces cerevisiae.” J Biol Chem. 2015 Mar 20;290(12):7517-34. doi: 10.1074/jbc.M114.633131. Epub 2015 Jan 28. PMID: 25631044
100. Andreyev AY, Tsui HS, Milne GL, Shmanai VV, Bekish AV, Fomich MA, Pham MN, Nong Y, Murphy AN, Clarke CF, Shchepinov MS. (2015) “Isotope-reinforced polyunsaturated fatty acids protect mitochondria from oxidative stress.” Free Radic Biol Med. 2015 May;82:63-72. doi: 10.1016/j.freeradbiomed.2014.12.023. Epub 2015 Jan 8. PMID: 25578654
99. Nguyen, T. P. T., Casarin, A., Desbats, M. A., Doimo, M., Trevisson, E., Santos-Ocana, C., Navas, P., Clarke, C. F., and Salviati, L. (2014) “Molecular characterization of the human COQ5 C-methyltransferase in Coenzyme Q10 biosynthesis. “Biochim Biophys Acta. 2014 Nov;1841(11):1628-38. doi: 10.1016/j.bbalip.2014.08.007. Epub 2014 Aug 23. PMID: 25152161
98. Shchepinov, M. S., Roginsky, V. A., Brenna, J. T., Molinari, R. J., To, R., Tsui, H., Clarke, C. F., and Manning-Bog, A. B. (2014) “Chapter 31 – Deuterium protection of polyunsaturated fatty acids against lipid peroxidation: A novel approach to mitigating mitochondrial neurological diseases”. In Omega-3 Fatty Acids in Brain and Neurological Health (Watson, ed.) pp. 373-383.
97. Chin, R.M., Fu, X., Pai, M.Y., Vergnes, L., Hwang, H., Deng, G., Diep, S., Lomenick, B., Meli, V.S., Monsalve, G.C., Hu, E., Whelan, S.A., Wang, J.X., Jung, G., Solis, G.M., Fazlollahi, F., Kaweeteerawat, C., Quach, A., Nili, M., Krall, A.S., Godwin, H.A., Chang, H.R., Faull, K.F., Guo, F., Jiang, M., Trauger, S.A., Saghatelian, A., Braas, D., Christofk, H.R, Clarke, C.F., Teitell, M.A., Petrascheck, M., Reue, K., Jung, M.E., Frand, A.R., Huang, J. “The metabolite α-ketoglutarate extends lifespan by inhibiting ATP synthase and TOR.” Nature. 2014 Jun 19;510(7505):397-401. PMCID: In Process.
96. Clarke CF, Rowat AC, Gober JW. “Osmotic stress: Is CoQ a membrane stabilizer?”. Nat Chem Biol. 2014 Apr;10(4):242-3. PMCID: In Process
95. He, C.H., Xie, L. X., Allan, C. M., Tran, U. C., and Clarke, C. F. (2014) “Coenzyme Q supplementation or over-expression of the yeast Coq8 kinase stabilizes multi-subunit Coq polypeptides complexes in yeast coq null mutants.” Biochim Biophys Acta 1841, 630-644. PMCID: PMC3959571.
94. Lamberson, C. R., Xu, L., Muchalski, H., Montenegro-Burke, J. R., Shmanai, V. V., Bekish, A. V., McLean, J. A., Clarke, C. F., Shchepinov, M. S., and Porter, N. A. (2014) “Unusual kinetic isotope effects of deuterium reinforced polyunsaturated fatty acids in tocopherol mediated free radical chain oxidations.” J. Amer. Chem. Soc. 136, 838-841. PMCID: In Process.
93. Ashraf, S., Gee, H. Y., Woerner, S., Xie, L. X., Vega-Warner, V., Svjetlana, L., Fang, H., Song, X., Cattran, D. C., Avila-Casado, C., Paterson, A. D., Nitschke, P., Bole-Feysot, C., Cochat, P., Esteve-Rudd, J., Haberberger, B., Allen, S. J., Zhou, W., Airik, R., Otto, E. A., Barua, M., Al-Hamed, M. H., Kari, J. A., Bockenhauer, D., Kleta, R., El Desoky, S., Hacihamdioglu, D. O., Gok, F., Washburn, J., Wiggins, R. C., Choi, M., Lifton, R. P., Levy, S., Han, Z., Salviati, L., Prokisch, H., Williams, D. S., Pollak, M., Clarke, C. F., Pei, Y., Antignac, C., and Hildebrandt, F. (2013) “ADCK4 mutations promote steroid-resistant nephrotic syndrome through CoQ10 biosynthesis disruption.” J. Clin. Invest. 123, 5179-5189. PMCID: PMC3859425.
92. Gasser, D. L., Winkler, C. A., Peng M., Ping, A., McKenzie, L. M., Kirk, G. D., Shi, Y., Xie, L. X., Marbois, B. N., Clarke, C. F., and Kopp, J. B. (2013) “Focal Segmental Glomerulosclerosis is associated with a PDSS2 haplotype and independently, with a decreased content of coenzyme Q10.” Amer. J. Physiol. Renal Physiol. Am J Physiol Renal Physiol. Oct; 305(8):F1228-38. PMCID: PMC3798722.
91. Allan, C. M., Hill, S., Morvaridi, S., Saiki, R., Johnson, J. S., Liau, W-S., Hirano, K., Kawashima, T., Ziming, J., Loo, J. A., Shepherd, J. N., and Clarke, C. F. (2013) “A conserved START domain coenzyme Q-binding polypeptide is required for efficient Q biosynthesis, respiratory electron transport, and antioxidant function in Saccharomyces cerevisiae“. Biochim. Biophys. Acta 1831, 776-791. PMCID: PMC3909687.
90. Gomez, F., Monsalve, G. C., Tse, V., Saiki, R., Weng, E., Lee, L., Srinivasan, C., Frand, A. R., and Clarke, C. F. (2012) “Delayed accumulation of intestinal coliform bacteria enhance life span and stress resistance in Caenorhabditis elegans fed respiratory deficient E. coli.“ BMC Microbiol. 12, 300. PMCID: PMC3548685.
89. Nguyen, T. P. T., Clarke, C. F. (2012) “Folate status of gut microbiome affects Caenorhabditis elegans lifespan“. BMC Biology 10, 66 PMCID: PMC3409036.
88. Gomez, F., Saiki, R., Chin, R., Srinivasan, C., and Clarke, C. F. (2012) “Restoring de novo coenzyme Q biosynthesis in Caenorhabditis elegans coq-3 mutants yields profound rescue compared to exogenous coenzyme Q supplementation“. Gene 506, 106-116. PMCID: PMC3437764.
87. Hill, S., Lamberson, C. R., Libin X., To, R., Tsui, H. S., Shmanai, V. V., Bekish, A. V., Awad, A. M., Marbois, B. N., Cantor, C. R., Porter, N. A., Clarke, C. F., and Shchepinov, M. S. (2012) “Small amounts of isotope-reinforced polyunsaturated fatty acids suppress lipid autoxidation.” Free Rad. Biol. Med. 53, 893-906. PMCID: PMC3437768.
86. Xie, L. X., Ozeir, M., Tang, J. Y., Chen, J. Y., Jaquinod, S-K., Fontecave, M, Clarke, C. F., and Pierrel, F. (2012) “Over-expression of the Coq8 kinase in Saccharomyces cerevisiae coq null mutants allows for accumulation of diagnostic intermediates of the coenzyme Q6 biosynthetic pathway.” J. Biol. Chem. 287, 23571-23581. PMCID: PMC3390632.
85. Rahman, S., Clarke, C. F., and Hirano, M. (2011) “176th ENMC International Workshop: Diagnosis and treatment of coenzyme Q10 deficiency”. Neuromuscular Disorders. 22, 76-86. PMCID: PMC3222743. REVIEW ARTICLE
84. Clarke, C. F. (2011) “Coq6 hydroxylase: Unmasked and bypassed.” Chem. Biol. 18, 1069-1070. PMCID:PMC3245979. INVITED COMMENTARY
83. Pfeiffer, M., Kayser, E.B., Yang, X., Abramson, E., Kenaston, M. A., Lago, C. U., Lo, H.H., Sedensky, M. M., Lunceford, A., Clarke, C. F., Wu, S. J., McLeod, C., Finkel, T., Morgan, P., Mills, E. M. (2011) “Caenorhabditis elegans UCP4 controls complex II-mediated oxidative phosphorylation through succinate transport.” J. Biol. Chem. 286, 37712-37720. PMCID:PMC3199514.
82. Falk, M. J., Polyak, E., Zhang, Z., Peng, M., King, R., Maltzman, J. S., Okwuego, E., Horyn, O., Nakamaru-Ogiso, E., Ostrovsky, J., Xie, L. X., Chen, J. Y., Marbois, B., Nissim, I., Clarke, C. F., Gasser, D. L. (2011) “Probucol ameliorates renal and metabolic sequelae of primary CoQ deficiency in Pdss2 mutant mice.” EMBO Mol. Med. 7, 410-427. PMCID: PMC3394513.
81. Heeringa, S. F., Chernin, G., Chaki, M., Zhou, W., Sloan, A. J., Ji, Z., Xie, L. X., Salviati, L., Hurd, T. W., Vega-Warner, V., Killen, P. D., Raphael Y., Ashraf, S., Ovunc, B., Schoeb, D. S., McLaughlin, H. M., Airik, R., Vlangos, C. N., Gbadegesin, R., Hinkes, B., Saisawat, P., Trevisson, E., Doimo, M., Casarin, A., Pertegato, V., Giorgi, G., Prokisch, H., Rötig, A., Antignac, C., Nurnberg, G., Becker, C., Wang, S., Ozaltin, F., Topaloglu, R., Bakkaloglu, A., Bakkaloglu, S. A., Müller, D., Beissert, A., Mir, S., Berdeli, A., Özen, S., Zenker, M., Verena Matejas, V., Ocana, C. S., Navas, P., Kusakabe, T., Kispert, A., Akman, S., Soliman, N. A., Krick, S., Mundel, P., Reiser J., Peter Nurnberg, P., Clarke, C. F., Wiggins, R. C., Faul, C., and Hildebrandt, F. (2011) “COQ6 mutations in human patients produce nephrotic syndrome with sensorineural deafness.” J. Clin. Invest. (2011) 121, 2013-2024. PMCID: PMC3083770.
80. Xie, L. X., Hsieh, E. J., Watanabe, S., Allan, C. M., Chen, J. Y., Tran, U. C., and Clarke, C. F. (2011) “Expression of the human atypical kinase ADCK3 rescues coenzyme Q biosynthesis and phosphorylation of Coq polypeptides in yeast coq8 mutants.” Biochim. Biophys. Acta (2011) 1811, 348-360. PMCID: PMC3075350.
79. Hill, S., Hirano, K., Shmanai, V. V., Marbois, B. N., Vidovic, D., Bekish, A. V., Kay, B., Tse, V., Fine, J., Clarke, C. F., and Shchepinov, M. S., (2011) “Isotope-reinforced polyunsaturated fatty acids strongly protect yeast cells from oxidative stress.” Free Radical Biol. Med. 50, 130-138. PMCID: PMC3014413.